To expand on this, we’re going to look at a few different models of how the spatial distribution of populations influences their divergence, and particularly how these factor into different processes of speciation.
What comes first, ecological or genetic divergence?
The order of these two processes have been in debate for some time, and different aspects of species and the environment can influence how (or if) these processes occur.
Different spatial models of speciation
Generally, when we consider the spatial models for speciation we divide these into distinct categories based on the physical distance of populations from one another. Although there is naturally a lot of grey area (as there is with almost everything in biological science), these broad concepts help us to define and determine how speciation is occurring in the wild.
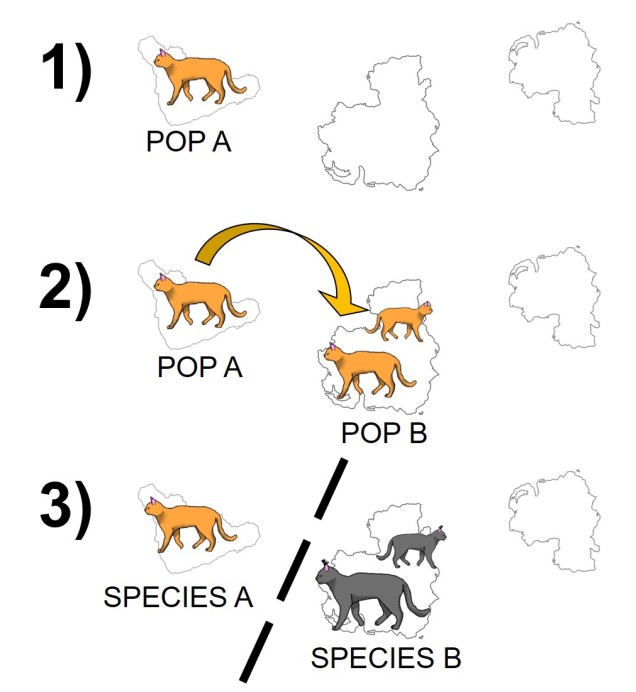
The standard model of allopatric speciation, following an island model. 1) We start with a single population occupying a single island. 2) A rare dispersal event pushes some individuals onto a new island, forming a second population. Note that this doesn’t happen often enough to allow for consistent gene flow (i.e. the island was only colonised once). 3) Over time, these populations may accumulate independent genetic and ecological changes due to both natural selection and drift, and when they become so different that they are reproductively isolated they can be considered separate species.
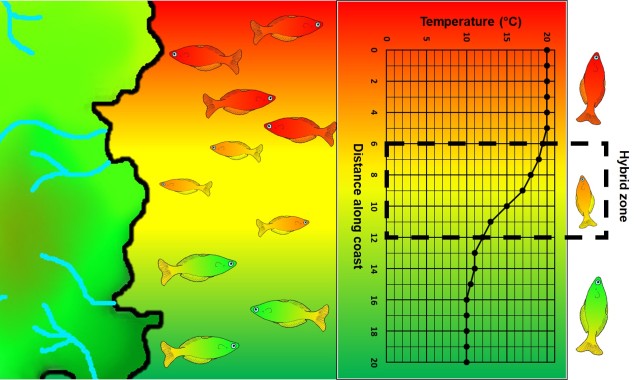
A step closer in bringing populations geographically together in speciation is “parapatry” and “peripatry”. Parapatric populations are often geographically close together but not overlapping: generally, the edges of their distributions are touching but do not overlap one another. A good analogy would be to think of countries that share a common border. Parapatry can occur when a species is distributed across a broad area, but some form of narrow barrier cleaves the distribution in two: this can be the case across particular environmental gradients where two extremes are preferred over the middle.
An example of parapatric species across an environment gradient (in this case, a temperature gradient along the ocean coastline). Left: We have two main species (red and green fish) which are adapted to either hotter or colder temperatures (red and green in the gradient), respectively. A small zone of overlap exists where hybrid fish (yellow) occur due to intermediate temperature. Right: How the temperature varies across the system, forming a steep gradient between hot and cold waters.
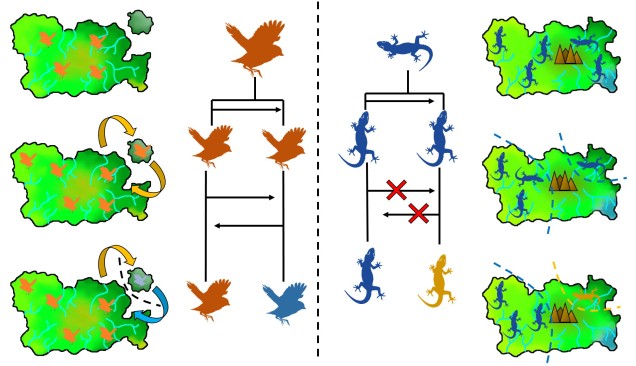
The two main ways peripatric species can form. Left: The dispersal method. In this example, there is a central ‘source’ population (orange birds on the main island), which holds most of the distribution. However, occasionally (more frequently than in the allopatric example above) birds can disperse over to the smaller island, forming a (mostly) independent secondary population. If the gene flow between this population and the central population doesn’t overwhelm the divergence between the two populations (due to selection and drift), then a new species (blue birds) can form despite the gene flow. Right: The range contraction method. In this example, we start with a single widespread population (blue lizards) which has a rapid reduction in its range. However, during this contraction one population is separated from the main body (i.e. as a refugia), which may also be a precursor of peripatric speciation.
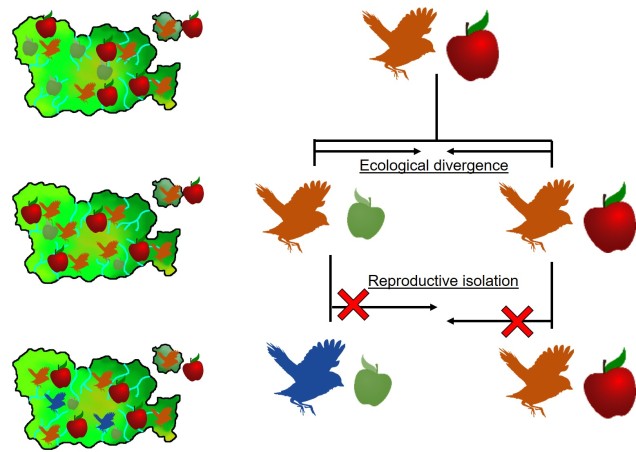
This can be tricky to visualise, so let’s invent an example. Say we have a tropical island, which is occupied by one bird species. This bird prefers to eat the large native fruit of the island, although there is another fruit tree which produces smaller fruits. However, there’s only so much space and eventually there are too many birds for the number of large fruit trees available. So, some birds are pushed to eat the smaller fruit, and adapt to a different diet, changing physiology over time to better acquire their new food and obtain nutrients. This shift in ecological niche causes the two populations to become genetically separated as small-fruit-eating-birds interact more with other small-fruit-eating-birds than large-fruit-eating-birds. Over time, these divergences in genetics and ecology causes the two populations to form reproductively isolated species despite occupying the same island.
A diagram of the ecological speciation example given above. Note that ecological divergence occurs first, with some birds of the original species shifting to the new food source (‘ecological niche’) which then leads to speciation. An important requirement for this is that gene flow is somehow (even if not totally) impeded by the ecological divergence: this could be due to birds preferring to mate exclusively with other birds that share the same food type; different breeding seasons associated with food resources; or other isolating mechanisms.
As you can see, the processes and context driving speciation are complex to unravel and many factors play a role in the transition from population to species. Understanding the factors that drive the formation of new species is critical to understanding not just how evolution works, but also in how new diversity is generated and maintained across the globe (and how that might change in the future).
A number of timesbefore on The G-CAT, we’ve discussed the idea of using the frequency of different genetic variants (alleles) within a particular population or species to test a number of different questions about evolution, ecology and conservation. These are all based on the central notion that certain forces of nature will alter the distribution and frequency of alleles within and across populations, and that these patterns are somewhat predictable in how they change.
One particular distinction we need to make early here is the difference between allele frequency and allele identity. In these analyses, often we are working with the same alleles (i.e. particular variants) across our populations, it’s just that each of these populations may possess these particular alleles in different frequencies. For example, one population may have an allele (let’s call it Allele A) very rarely – maybe only 10% of individuals in that population possess it – but in another population it’s very common and perhaps 80% of individuals have it. This is a different level of differentiation than comparing how different alleles mutate (as in the coalescent) or how these mutations accumulate over time (like in many phylogenetic-based analyses).
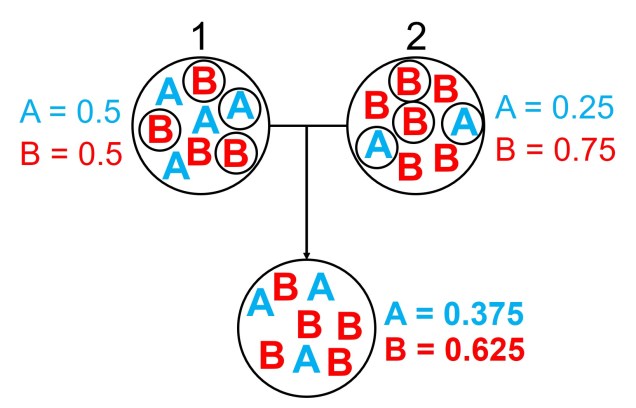
An example of the difference between allele frequency and identity.In this example (and many of the figures that follow in this post), the circle denote different populations, within which there are individuals which possess either an A gene (blue) or a B gene. Left: If we compared Populations 1 and 2, we can see that they both have A and B alleles. However, these alleles vary in their frequency within each population, with an equal balance of A and B in Pop 1 and a much higher frequency of B in Pop 2. Right: However, when we compared Pop 3 and 4, we can see that not only do they vary in frequencies, they vary in the presence of alleles, with one allele in each population but not the other.
An example of how gene flow across populations homogenises allele frequencies. We start with two initial populations (1 and 2 from above), which have very different allele frequencies. Hybridising individuals across the two populations means some alleles move from Pop 1 and Pop 2 into the hybrid population: which alleles moves is random (the smaller circles). Because of this, the resultant hybrid population has an allele frequency somewhere in between the two source populations: think of like mixing red and blue cordial and getting a purple drink.
An example of a Structure plot which long-term The G-CAT readers may be familiar with. This is taken from Brauer et al. (2013), where the authors studied the population structure of the Yarra pygmy perch. Each small column represents a single individual, with the colours representing how well the alleles of that individual fit a particular genetic population (each population has one colour). The numbers and broader columns refer to different ‘localities’ (different from populations) where individuals were sourced. This shows clear strong population structure across the 4 main groups, except for in Locality 6 where there is a mixture of Eastern and Merri/Curdies alleles.
Determining genetic bottlenecks and demographic change
A diagram of how allele frequencies change in genetic bottlenecks due to genetic drift. Left: Large circles again denote a population (although across different sequential times), with smaller circle denoting which alleles survive into the next generation (indicated by the coloured arrows). We start with an initial ‘large’ population of 8, which is reduced down to 4 and 2 in respective future times. Each time the population contracts, only a select number of alleles (or individuals) ‘survive’: assuming no natural selection is in process, this is totally random from the available gene pool. Right: We can see that over time, the frequencies of alleles A and B shift dramatically, leading to the ‘extinction’ of Allele B due to genetic drift. This is because it is the less frequent allele of the two, and in the smaller population size has much less chance of randomly ‘surviving’ the purge of the genetic bottleneck.
An example of how the frequency of alleles might vary under natural selection in correlation to the environment. In this example, the blue allele A is adaptive and under positive selection in the more intense environment, and thus increases in frequency at higher values. Contrastingly, the red allele B is maladaptive in these environments and decreases in frequency. For comparison, the black allele shows how the frequency of a neutral (non-adaptive or maladaptive) allele doesn’t vary with the environment, as it plays no role in natural selection.
Fixed differences are sometimes used as a type of diagnostic trait for species. This means that each ‘species’ has genetic variants that are not shared at all with its closest relative species, and that these variants are so strongly under selection that there is no diversity at those loci. Often, fixed differences are considered a level above populations that differ by allelic frequency only as these alleles are considered ‘diagnostic’ for each species.
An example of the difference between fixed differences and allelic frequency differences. In this example, we have 5 cats from 3 different species, sequencing a particular target gene. Within this gene, there are three possible alleles: T, A or G respectively. You’ll quickly notice that the T allele is both unique to Species A and is present in all cats of that species (i.e. is fixed). This is a fixed difference between Species A and the other two. Alleles A and G, however, are present in both Species B and C, and thus are not fixed differences even if they have different frequencies.
To distinguish between the two, we often use the overall frequency of alleles in a population as a basis for determining how likely two individuals share an allele by random chance. If alleles which are relatively rare in the overall population are shared by two individuals, we expect that this similarity is due to family structure rather than population history. By factoring this into our relatedness estimates we can get a more accurate overview of how likely two individuals are to be related using genetic information.
The wild world of allele frequency
Despite appearances, this is just a brief foray into the many applications of allele frequency data in evolution, ecology and conservation studies. There are a plethora of different programs and methods that can utilise this information to address a variety of scientific questions and refine our investigations.
Meaning: Cinis: from [ash] in Latin; descendens from [descends] in Latin.
Translation: descending from the ash; describes hunting behaviour in ash mountains of Vvardenfell.
Common name
Cliff racer
A cliff racer hovering above a precipice on Vvardenfell.
Taxonomic status
Kingdom Animalia; Phylum Chordata; Class Aves; Subclass Archaeornithes; Family Vvardidae; GenusCinis; Speciesdescendens
Conservation status
Least Concern [circa 3E 427]
Threatened [circa 4E 433]
Distribution
Once widespread throughout the north eastern region of Tamriel, occupying regions from the island of Vvardenfell to mainland Morrowind and Solstheim. Despite their name, the cliff racer is found across nearly all geographic regions of Vvardenfell, although the species is found in greatest densities in the rocky interior region of Stonefalls.
Following a purge of the species as part of pest control management, the cliff racer was effectively exterminated from parts of its range, including local extinction on the island of Solstheim. Since the cull the cliff racer is much less abundant throughout its range although still distributed throughout much of Vvardenfell and mainland Morrowind.
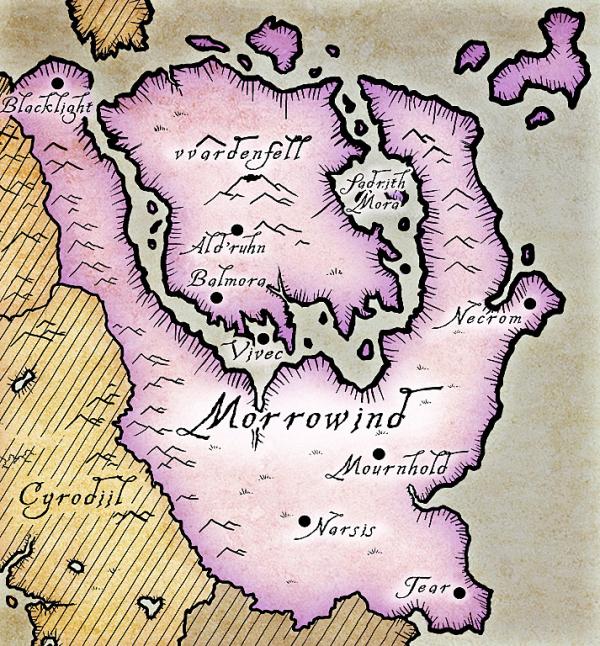
The province of Morrowind, which largely contains the distribution of the cliff racer. The island of Solstheim is found to the northwest of the map (the lower half of the island can be seen in brown).
Habitat
Although, much as the name suggests, the cliff racer prefers rocky outcroppings and mountainous regions in which it can build its nest, the species is frequently seen in lowland swamp and plains regions of Morrowind.
Behaviour and ecology
The cliff racer is a highly aggressive ambush predator, using height and range to descend on unsuspecting victims and lashing at them with its long, sharp tail. Although preferring to predate on small rodents and insects (such as kwama), cliff racers have been known to attack much larger beasts such as agouti and guar if provoked or desperate. The highly territorial nature of cliff racer means that they often attack travellers, even if they pose no immediate threat or have done nothing to provoke the animal.
A cliff racer descends upon its prey.
Despite the territoriality of cliff racers, large flocks of them can often be found in the higher altitude regions of Vvardenfell, perhaps facilitated by an abundance of food (reducing competition) or communal breeding grounds. Attempts by researchers to study these aggregations have been limited due to constant attacks and damage to equipment by the flock.
Following the control measures implemented, the population size of these populations of cliff racers declined severely; however, given the survival of the majority of the population it does not appear this bottleneck has severely impacted the longevity of the species. The extirpation of the Solstheim population of cliff racers likely removed a unique ESU from the species, given the relative isolation of the island. Whether the island will be recolonised in time by Vvardenfell cliff racers is unknown, although the presence of any cliff racers back onto Solstheim would likely be met with strong opposition from the local peoples.
Adaptive traits
The broad wings, dorsal sail and long tail allow the cliff racer to travel large distances in the air, serving them well in hunting behaviour. The drawback of this is that, if hunting during the middle hours of the day, the cliff racer leaves an imposing shadow on the ground and silhouette in the sky, often alerting aware prey to their presence. That said, the speed of descent and disorienting cry of the animal often startles prey long enough for the cliff racer to attack.
The plumes of the cliff racer are a well-sought-after commodity by local peoples, used in the creation of garments and household items. Whether these plumes serve any adaptive purpose (such as sexual selection through mate signalling) is unknown, given the difficulties with studying wild cliff racer behaviour.
Management actions
Although suffering from a strong population bottleneck after the purge, the cliff racer is still relatively abundant across much of its range and maintains somewhat stable size. Management and population control of the cliff racer is necessary across the full distribution of the species to prevent strong recovery and maintain public safety and ecosystem balance. Breeding or rescuing cliff racers is strictly forbidden and the species has been widely declared as ‘native pest’, despite the somewhat oxymoron nature of the phrase.