To expand on this, we’re going to look at a few different models of how the spatial distribution of populations influences their divergence, and particularly how these factor into different processes of speciation.
What comes first, ecological or genetic divergence?
The order of these two processes have been in debate for some time, and different aspects of species and the environment can influence how (or if) these processes occur.
Different spatial models of speciation
Generally, when we consider the spatial models for speciation we divide these into distinct categories based on the physical distance of populations from one another. Although there is naturally a lot of grey area (as there is with almost everything in biological science), these broad concepts help us to define and determine how speciation is occurring in the wild.
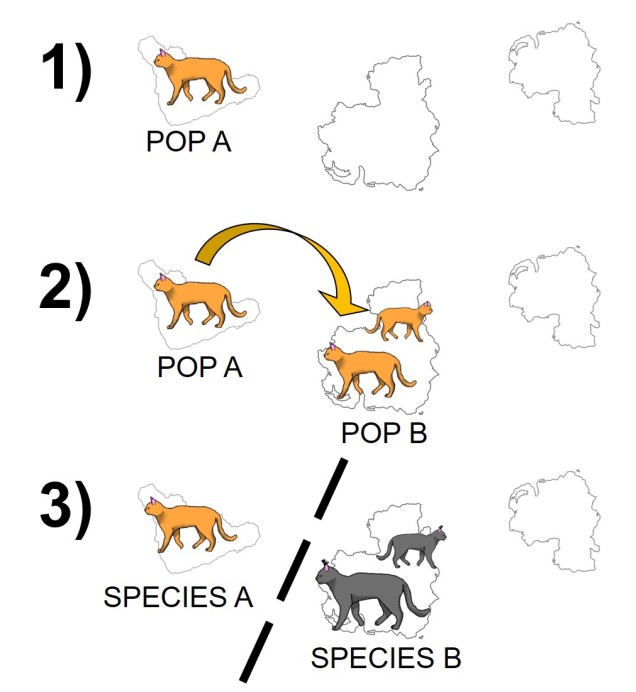
The standard model of allopatric speciation, following an island model. 1) We start with a single population occupying a single island. 2) A rare dispersal event pushes some individuals onto a new island, forming a second population. Note that this doesn’t happen often enough to allow for consistent gene flow (i.e. the island was only colonised once). 3) Over time, these populations may accumulate independent genetic and ecological changes due to both natural selection and drift, and when they become so different that they are reproductively isolated they can be considered separate species.
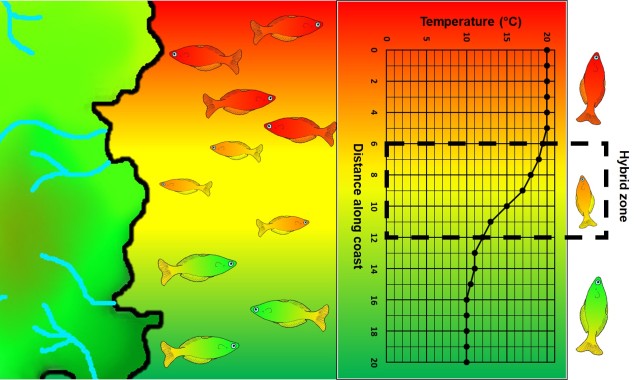
A step closer in bringing populations geographically together in speciation is “parapatry” and “peripatry”. Parapatric populations are often geographically close together but not overlapping: generally, the edges of their distributions are touching but do not overlap one another. A good analogy would be to think of countries that share a common border. Parapatry can occur when a species is distributed across a broad area, but some form of narrow barrier cleaves the distribution in two: this can be the case across particular environmental gradients where two extremes are preferred over the middle.
An example of parapatric species across an environment gradient (in this case, a temperature gradient along the ocean coastline). Left: We have two main species (red and green fish) which are adapted to either hotter or colder temperatures (red and green in the gradient), respectively. A small zone of overlap exists where hybrid fish (yellow) occur due to intermediate temperature. Right: How the temperature varies across the system, forming a steep gradient between hot and cold waters.
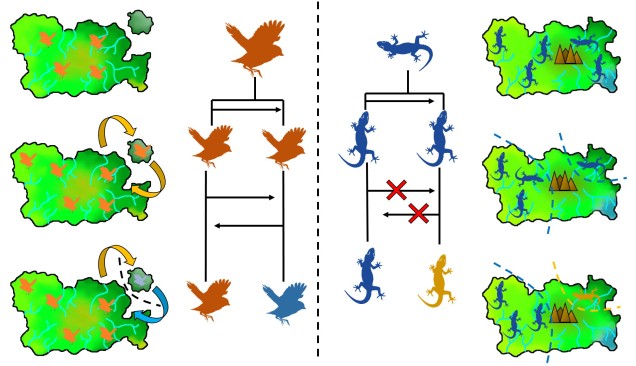
The two main ways peripatric species can form. Left: The dispersal method. In this example, there is a central ‘source’ population (orange birds on the main island), which holds most of the distribution. However, occasionally (more frequently than in the allopatric example above) birds can disperse over to the smaller island, forming a (mostly) independent secondary population. If the gene flow between this population and the central population doesn’t overwhelm the divergence between the two populations (due to selection and drift), then a new species (blue birds) can form despite the gene flow. Right: The range contraction method. In this example, we start with a single widespread population (blue lizards) which has a rapid reduction in its range. However, during this contraction one population is separated from the main body (i.e. as a refugia), which may also be a precursor of peripatric speciation.
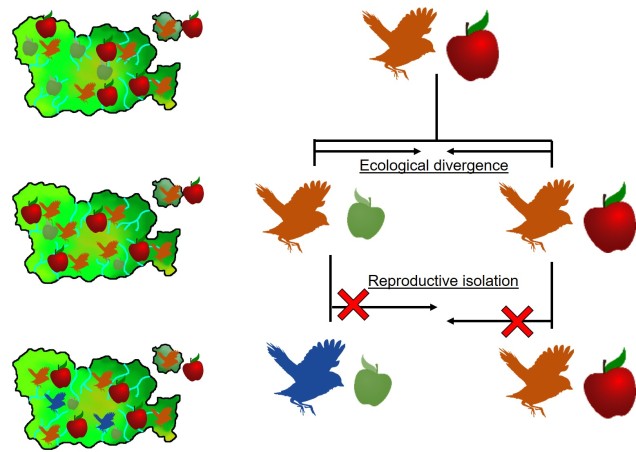
This can be tricky to visualise, so let’s invent an example. Say we have a tropical island, which is occupied by one bird species. This bird prefers to eat the large native fruit of the island, although there is another fruit tree which produces smaller fruits. However, there’s only so much space and eventually there are too many birds for the number of large fruit trees available. So, some birds are pushed to eat the smaller fruit, and adapt to a different diet, changing physiology over time to better acquire their new food and obtain nutrients. This shift in ecological niche causes the two populations to become genetically separated as small-fruit-eating-birds interact more with other small-fruit-eating-birds than large-fruit-eating-birds. Over time, these divergences in genetics and ecology causes the two populations to form reproductively isolated species despite occupying the same island.
A diagram of the ecological speciation example given above. Note that ecological divergence occurs first, with some birds of the original species shifting to the new food source (‘ecological niche’) which then leads to speciation. An important requirement for this is that gene flow is somehow (even if not totally) impeded by the ecological divergence: this could be due to birds preferring to mate exclusively with other birds that share the same food type; different breeding seasons associated with food resources; or other isolating mechanisms.
As you can see, the processes and context driving speciation are complex to unravel and many factors play a role in the transition from population to species. Understanding the factors that drive the formation of new species is critical to understanding not just how evolution works, but also in how new diversity is generated and maintained across the globe (and how that might change in the future).
Although we often focus on the genetic traits of species in molecular ecology studies, the physiological (or phenotypic) traits are equally as important in shaping their evolution. These different traits are not only the result themselves of evolutionary forces but may further drive and shape evolution into the future by changing how an organism interacts with the environment.
There are a massive number of potential traits we could focus on, each of which could have a large number of different (and interacting) impacts on evolution. One that is often considered, and highly relevant for genetic studies, is the influence of dispersal capability.
Dispersal
Dispersal is essentially the process of an organism migrating to a new habitat, to the point of the two being used almost interchangeably. Often, however, we regard dispersal as a migration event that actually has genetic consequences; particularly, if new populations are formed or if organisms move from one population to another. This can differ from straight migration in that animals that migrate might not necessarily breed (and thus pass on genes) into a new region during their migration; thus, evidence of those organisms will not genetically proliferate into the future through offspring.
Naturally, the ability of organisms to disperse is highly variable across the tree of life and reliant on a number of other physiological factors. Marine mammals, for example, can disperse extremely far throughout their lifetimes, whereas some very localised species like some insects may not move very far within their lifetime at all. The movement of organisms directly facilitates the movement of genetic material, and thus has significant impacts on the evolution and genetic diversity of species and populations.
The (simplistic) relationship between dispersal capability and one aspect of population genetics, population structure (measured as Fst). As organisms are more capable of dispersing longer distance (or more frequently), the barriers between populations become weaker.
As these individuals occupy large ranges, localised impacts are unlikely to critically affect their full distribution. Individual organisms that are occupying an unpleasant space can easily move to a more favourable habitat (provided that one exists). Furthermore, with a large population (which is more likely with highly dispersive species), genetic drift is substantially weaker and natural selection (generally) has a higher amount of genetic diversity to work with. This is, of course, assuming that dispersal leads to a large overall population, which might not be the case for species that are critically endangered (such as the cheetah).
The Wright island model of population structure. In this example, different independent populations are labelled in the bold letters, with dispersal pathways demonstrated by the different arrows. In the island model, dispersal is equally likely between all populations (including from B–D in this example, even though there aren’t any arrows showing it). Naturally, this is not overly realistic and so the island model is used mostly as a neutral, base model.
Intermediately dispersing species
A large number of species, however, are likely to occupy a more intermediate range of dispersal ability. These species might be able to migrate to neighbouring populations, or across a large proportion of their geographic range, but individuals from one end of the range are still somewhat isolated from individuals at the other end.
This often leads to some effect of population structure; different portions of the geographic range are genetically segregated from one another depending on how much gene flow (i.e. dispersal) occurs between populations. In the most simplest scenario, this can lead to what we call isolation-by-distance. Rather than forming totally independent populations, gene flow occurs across short ranges between adjacent ‘populations’. This causes a gradient of genetic differentiation, with one end of the range being clearly genetically different to the other end, with a gradual slope throughout the range. We see this often in marine invertebrates, for example, which might have somewhat localised dispersal but still occupy a large range by following oceanographic currents.
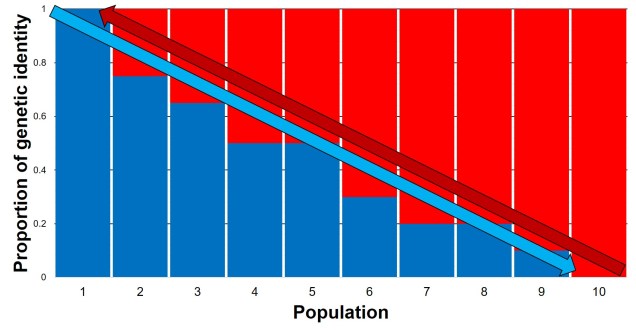
An example of how an isolation-by-distance population network might come about. In this example, we have a series of populations (the different pie charts) spread throughout a river system (that blue thing). The different pie charts represent how much of the genetics of that population matches one end of the river: either the blue end (left) or red end (right). Populations can easily disperse into adjacent populations (the green arrows) but less so to further populations. This leads to gradual changes across the length of the river, with the far ends of the river clearly genetically distinct from the opposite end but relatively similar to neighbouring populations.The genetic representation of the above isolation-by-distance example. Each column represents a single population (in the previous figure, a pie chart), with the different colours also representing the relative genetic identity of that population. As you can see, moving from Population 1 to 10 leads to a gradient (decreasing) in blue genes but increase in red genes. The inverse can be said moving in the opposite direction. That said, comparing Population 1 and Population 10 shows that they’re clearly different, although there is no clear cut-off point across the range of other populations.
Medium dispersal capabilities are also often a requirement for forming ‘metapopulations’. In this population arrangement, several semi-independent populations are present within the geographic range of the species. Each of these are subject to their own local environmental pressures and demographic dynamics, and because of this may go locally extinct at any given time. However, dispersal connections between many of these populations leads to recolonization and gene flow patterns, allowing for extinction-dispersal dynamics to sustain the overall metapopulation. Generally, this would require greater levels of dispersal than those typically found within metapopulation species, as individuals must traverse uninhabitable regions relatively frequently to recolonise locally extinct habitat.
An example of metapopulation dynamics. Different subpopulations (lettered circles) are connected via dispersal (arrows). These different subpopulations can be different sizes and are mostly independent of one another, meaning that a single subpopulation can go locally extinct (the red X) without collapsing the entire system. The different dispersal pathways mean that one population can recolonise extinct habitat and essentially ‘rebirth’ other subpopulations (the green arrows).
Species with low dispersal capabilities are often at risk of local extinction and are unable to easily recolonise these habitats after the event has ended. Their movement is often restricted to rare environmental events such as flooding that carry individuals long distances despite their physiological limitations. Because of this, low dispersal species are often at greater risk of total extinction and extinction vertices than their higher dispersing counterparts.
Accounting for dispersal in population genetics
Incorporating biological and physiological aspects of our study taxa is important for interpreting the evolutionary context of species. Dispersal ability is but one of many characteristics that can influence the ability of species to respond to selective pressures, and the context in which this natural selection occurs. Thus, understanding all aspects of an organism is important in building the full picture of their evolution and future prospects.
A map representing the relative proportion of endemic species in Australia, generated through the Atlas of Living Australia. The colours range from no (white; 0% endemics) or little (blue) to high levels of endemism (red; 100% of species are endemic). As you can see, some biogeographic hotspots are clearly indicated (southwest WA, the east coast, the Kimberley ranges).
But the real question is: why are there so many endemics in Australia? What is so special about our country that lends to our unique flora and fauna? Although we naturally associate tropical regions with lush, vibrant and diverse life, most of Australia is complete desert. That said, most of our species are concentrated in the tropical regions of the country, particularly in the upper east coast and far north (the ‘Top End’).
There are a number of different factors which contribute to the high species diversity of Australia. Most notably is how isolated we are as a continent: Australia has been separated from most of the rest of the world for millions of years. In this time, the climate has varied dramatically as the island shifted northward, creating a variety of changing environments and unique ecological niches for species to specialise into. We refer to these species groups as ‘Gondwana relicts’, since their last ancestor with the rest of the world would have been distributed across the supercontinent Gondwana over 100 million years ago. These include marsupials, many birds groups (including ratites and megapodes), many fish groups and a plethora of others. A Gondwanan origin explains why they are only found within Australia, southern Africa and South America (the closest landmass that was also historically connected to Gondwana).
Early arrivals and naturalisation to the Australian ecosystem
Eventually, this connection also brought with them one of our most iconic species; the dingo. Estimates of their arrival dates the migration at around 6 thousand years ago. As Australia’s only ‘native’ dog, there has been much debate about its status as an Australian icon. To call the dingo ‘native’ implies it’s always been there: but 6 thousand years is more than enough time to become ingrained within the ecosystem in a stable fashion. So, to balance the debate (and prevent the dingo from being labelled as an ‘invasive pest’ unfairly), we often refer to them as ‘naturalised’. This term helps us to disentangle modern-day pests, many of which our immensely destructive to the natural environment, from other species that have naturally migrated and integrated many years ago.
Although it may not be a “true native”, the dingo will forever be a badge of our native species pride.
Invaders of the Australian continent
Of course, we can never ignore the direct impacts of humans on the ecosystem. Particularly with European settlement, another plethora of animals were introduced for the first time into Australia; these were predominantly livestock animals or hunting-related species (both as predators and prey). This includes the cane toad, widely regarded as one of the biggest errors in pest control on the planet.
When European settlers in the 1930s attempted to grow sugar cane in the far eastern part of the country, they found their crops decimated by a local beetle. In an effort to eradicate them, they brought over a species of cane toad, with the idea that they would control the beetle population and all would be well. Only, cane toads are particularly lazy and instead of targeting the cane beetles, they just thrived on all the other native invertebrates around. They’re also very resilient and adaptable (and highly toxic), so their numbers exploded and they’ve since spread across a large swathe of the country. Their toxic skin makes them fatal food objects for many native predators and they strongly compete against other similar native animals (such as our own amphibians). The cane toad introduction of 1935 is the poster child of how bad failed pest control can be.
This guy here, he’s a bastard. Spotted in my parent’s backyard in Ipswich, QLD. Source: me, with spite.
But is native always better?
History tells a very stark tale about the poor native animals and the ravenous, rampaging pest species. Because of this, it is a widely adopted philosophical viewpoint that ‘native is always best’. And while I don’t disagree with the sentiment (of course we need to preserve our native wildlife, and not the massively overabundant pests), there are rare examples where nature is a little more complicated. In Australia, this is exemplified in the noisy miner.
The noisy miner is a small bird which, much like its name implies, is incredibly noisy and aggressive. It’s highly abundant, found predominantly throughout urban and suburban areas, and seems to dominate the habitat. It does this by bullying out other bird species from nesting grounds, creating a monopoly on the resource to the exclusion of many other species (even larger ones such as crows and magpies). Despite being native, it seems to have thrived on human alteration of the landscape and is a serious threat to the survival and longevity of many other species. If we thought of it solely under the ‘nature is best’ paradigm, we would dismiss the noisy miner as ‘doing what it should be.’ The truth is really more of a philosophical debate: is it natural to let the noisy miner outcompete many other natives, possibly resulting in their extinction? Or is it only because of human interference (and thus is our responsibility to fix) that the noisy miner is doing so well in the first place? It’s not a simple question to answer, although the latter seems to be incredibly important.
An example of the aggressive behaviour of the noisy miner (top), swooping down on a pied currawong (bottom). Despite the size differences, noisy miners will frequently attempt to harass and scare off other larger birds. Image source: Bird Ecology Study Group website.
The amazing biodiversity of Australia is a badge of honour we should wear with patriotic pride. Conservation efforts of our endemic fauna are severely limited by a lack of funding and resources, and despite a general acceptance of the importance of diverse ecosystems we remain relatively ineffective at preserving it. Understanding and connecting with our native wildlife, whilst finding methods to control invasive species, is key to conserving our wonderful ecosystems.
The distribution of organisms across the Earth, both over time and across space, is a fundamental aspect of the field of biogeography. But our understanding of the mechanisms by which organisms are distributed across the globe, and how this affects their evolution, can be at times highly enigmatic. Why are Australia and the Americas the only two places that have marsupials? How did lemurs get all the way to Madagascar, and why are they the only primate that has made the trip? How did Darwin’s famous finches get over to the Galápagos, and why are there so many species of them there now?
All of these questions can be addressed with a combination of genetic, environmental and ecological information across a variety of timescales. However, the overall field of biogeography (and phylogeography as a derivative of it) has traditionally been largely rooted on a strong yet changing theoretical basis. The earliest discussions and discoveries related to biogeography as a field of science date back to the 18th Century, and to Carl Linnaeus (to whom we owe our binomial classification system) and Alexander von Humboldt. These scientists (and undoubtedly many others of that era) were among the first to notice how organisms in similar climates (e.g. Australia, South Africa and South America) showed similar physical characteristics despite being so distantly separated (both in their groups and geographic distance). The communities of these regions also appeared to be highly similar. So how could this be possible over such huge distances?
A pretty unreasonable mechanism (and example) of dispersal in foxes. And yes, all tourists wear sunglasses and Hawaiian shirts, even arctic fox ones.
Dispersal or vicariance?
Two main explanations for these patterns are possible; dispersal and vicariance. As one might expect, dispersal denotes that an ancestral species was distributed in one of these places (referred to as the ‘centre of origin’) before it migrated and inhabited the other places. Contrastingly, vicariancesuggests that the ancestral species was distributed everywhere originally, covering all contemporary ranges within it. However, changes in geography, climate or the formation of other barriers caused the range of the ancestor to fragment, with each fragmented group evolving into its own distinct species (or group of species).
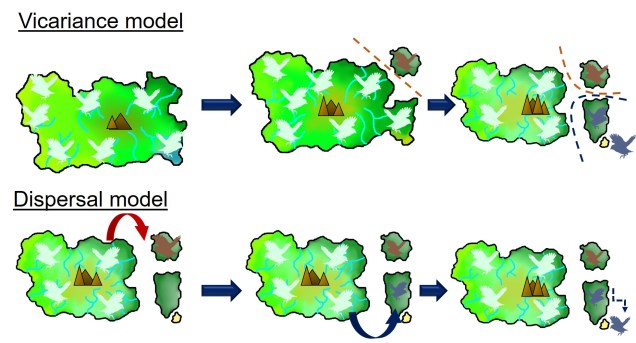
An example of dispersal vs. vicariance patterns of biogeography in an island bird (pale blue). In the top example, the sequential separation of parts of the island also cause parts of the distribution of the original bird species to become fragmented. These fragments each evolve independently of their ancestor and form new species (red, and then blue). In the bottom example, the island geography doesn’t change but in rare events a bird disperses from the main island onto a new island. The new selective pressures of that island cause the dispersed birds to evolve into new species (red and blue). In both examples, islands that were recently connected or are easy to disperse across do not generate new species (in the sandy island in the bottom right). You’ll notice that both processes result in the same biogeographic distribution of species.
In initial biogeographic science, dispersal was the most heavily favoured explanation. At the time, there was no clear mechanism by which organisms could be present all over the globe without some form of dispersal: it was generally believed that the world was a static, unmoving system. Dispersal was well supported by some biological evidence such as the diversification of Darwin’s finches across the Galápagos archipelago. Thus, this concept was supported through the proposals of a number of prominent scientists such as Charles Darwin and A.R. Wallace. For others, however, the distance required for dispersal (such as across entire oceans) seemed implausible and biologically unrealistic.
A paradigm shift in biogeography
Two particular developments in theory are credited with a paradigm shift in the field; cladistics and plate tectonics. Cladistics simply involved using shared biological characteristics to reconstruct the evolutionary relationships of species (think like phylogenetics, but using physical traits instead of genetic sequence). Just as importantly, however, was plate tectonic theory, which provided a clear way for organisms to spread across the planet. By understanding that, deep in the past, all continents had been directly connected to one another provides a convenient explanation for how species groups spread. Instead of requiring for species to travel across entire oceans, continental drift meant that one widespread and ancient ancestor on the historic supercontinent (Pangaea; or subsequently Gondwana and Laurasia) could become fragmented. It only required that groups were very old, but not necessarily very dispersive.
Surf’s up, dudes! Although continental drift was no doubt an important factor in the distribution and dispersal of many organisms on Earth, it actually probably wasn’t the reason lemurs got to Madagascar. Sorry for the mislead.
From these advances in theory, cladistic vicariance biogeography was born. The field rapidly overtook dispersal as the most likely explanation for biogeographic patterns across the globe by not only providing a clear mechanism to explain these but also an analytical framework to test questions relating to these patterns. Further developments into the analytical backbone of cladistic vicariance allowed for more nuanced questions of biogeography to be asked, although still fundamentally ignored the role of potential dispersals in explaining species’ distributions.
Modern philosophy of biogeography
So, what is the current state of the field? Well, the more we research biogeographic patterns with better data (such as with genomics) the more we realise just how complicated the history of life on Earth can be. Complex modelling (such as Bayesian methods) allow us to more explicitly test the impact of Earth history events on our study species, and can provide more detailed overview of the evolutionary history of the species (such as by directly estimating times of divergence, amount of dispersal, extent of range shifts).
From a theoretical perspective, the consistency of patterns of groups is always in question and exactly what determines what species occurs where is still somewhat debatable. However, the greater number of types of data we can now include (such as geological, paleontological, climatic, hydrological, genetic…the list goes on!) allows us to paint a better picture of life on Earth. By combining information about what we know happened on Earth, with what we know has happened to species, we can start to make links between Earth history and species history to better understand how (or if) these events have shaped evolution.