It should come as no surprise to any reader of The G-CAT that I’m a firm believer against the false dichotomy (and yes, I really do love that phrase) of “nature versus nurture.” Primarily, this is because the phrase gives the impression of some kind of counteracting balance between intrinsic (i.e. usually genetic) and extrinsic (i.e. usually environmental) factors and how they play a role in behaviour, ecology and evolution. While both are undoubtedly critical for adaptation by natural selection, posing this as a black-and-white split removes the possibility of interactive traits.
The real Circle of Life. Not only do genes and the environment interact with one another, but genes may interact with other genes and environments may be complex and multi-faceted.
A very simplified example of adaptation from genetic variation. In this example, we have two different alleles of a single gene (orange and blue). Natural selection favours the blue allele so over time it increases in frequency. The difference between these two alleles is at least one base pair of DNA sequence; this often arises by mutation processes.
Despite how important the underlying genes are for the formation of proteins and definition of physiology, they are not omnipotent in that regard. In fact, many other factors can influence how genetic traits relate to phenotypic traits: we’ve discussed a number of these in minor detail previously. An example includes interactions across different genes: these can be due to physiological traits encoded by the cumulative presence and nature of many loci (as in quantitative trait loci and polygenic adaptation). Alternatively, one gene may translate to multiple different physiological characters if it shows pleiotropy.
Differential expression
One non-direct way genetic information can impact on the phenotype of an organism is through something we’ve briefly discussed before known as differential expression. This is based on the notion that different environmental pressures may affect the expression (that is, how a gene is translated into a protein) in alternative ways. This is a fundamental underpinning of what we call phenotypic plasticity: the concept that despite having the exact same (or very similar) genes and alleles, two clonal individuals can vary in different traits. The is related to the example of genetically-identical twins which are not necessarily physically identical; this could be due to environmental constraints on growth, behaviour or personality.
An example of differential expression in wild populations of southern pygmy perch, courtesy of Brauer et al. (2017). In this figure, each column represents a single individual fish, with the phylogenetic tree and coloured boxes at the top indicating the different populations. Each row represents a different gene (this is a subset of 50 from a much larger dataset). The colour of each cell indicates whether the expression of that gene is expressed more (red) or less (blue) than average. As you can see, the different populations can clearly be seen within their expression profiles, with certain genes expressing more or less in certain populations.
The discovery of epigenetic markers and their influence on gene expression has opened up the possibility of understanding heritable traits which don’t appear to be clearly determined by genetics alone. For example, research into epigenetics suggest that heritable major depressive disorder (MDD) may be controlled by the expression of genes, rather than from specific alleles or genetic variants themselves. This is likely true for a number of traits for which the association to genotype is not entirely clear.
Epigenetic adaptation?
From an evolutionary standpoint again, epigenetics can similarly influence the ‘bang for a buck’ of particular genes. Being able to translate a single gene into many different forms, and for this to be linked to environmental conditions, allows organisms to adapt to a variety of new circumstances without the need for specific adaptive genes to be available. Following this logic, epigenetic variation might be critically important for species with naturally (or unnaturally) low genetic diversity to adapt into the future and survive in an ever-changing world. Thus, epigenetic information might paint a more optimistic outlook for the future: although genetic variation is, without a doubt, one of the most fundamental aspects of adaptability, even horrendously genetically depleted populations and species might still be able to be saved with the right epigenetic diversity.
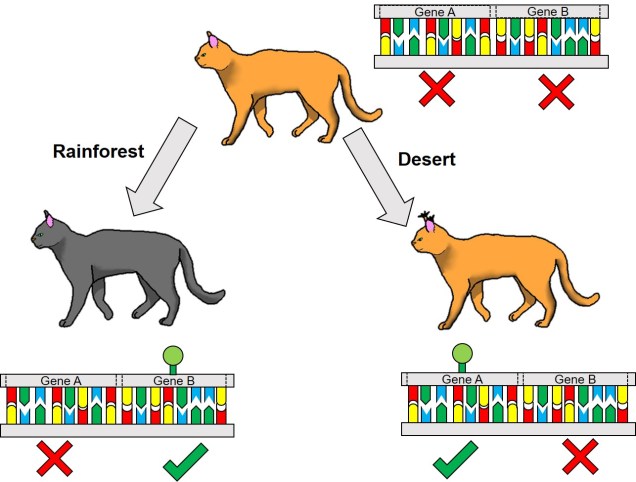
A relatively simplified example of adaptation from epigenetic variation. In this example, we have a species of cat; the ‘default’ cat has non-tufted ears and an orange coat. These two traits are controlled by the expression of Genes A and B, respectively: in the top cat, neither gene is expressed. However, when this cat is placed into different environments, the different genes are “switched on” by epigenetic factors (the green markers). In a rainforest environment, the dark foliage makes darker coat colour more adaptive; switching on Gene B allows this to happen. Conversely, in a desert environment switching on Gene A causes the cat to develop tufts on its ears, which makes it more effective at hunting prey hiding in the sands. Note that in both circumstances, the underlying genetic sequence (indicated by the colours in the DNA) is identical: only the expression of those genes change.
Epigenetic research, especially from an ecological/evolutionary perspective, is a very new field. Our understanding of how epigenetic factors translate into adaptability, the relative performance of epigenetic vs. genetic diversity in driving adaptability, and how limited heritability plays a role in adaptation is currently limited. As with many avenues of research, further studies in different contexts, experiments and scopes will reveal further this exciting new aspect of evolutionary and conservation genetics. In short: watch this space! And remember, ‘nature is nurture’ (and vice versa)!
There are quite literally millions of species on Earth, ranging from the smallest of microbes to the largest of mammals. In fact, there are so many that we don’t actually have a good count on the sheer number of species and can only estimate it based on the species we actually know about. Unsurprisingly, then, the number of species vastly outweighs the number of people that research them, especially considering the sheer volumes of different aspects of species, evolution, conservation and their changes we could possibly study.
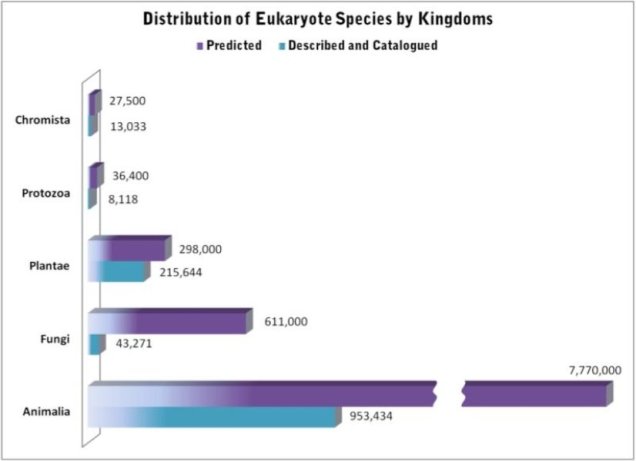
Some estimations on the number of eukaryotic species (i.e. not including things like bacteria), with the number of known species in blue and the predicted number of total species on Earth in purple. Source: Census of Marine Life.
This is partly where the concept of a ‘model’ comes into it: it’s much easier to pick a particular species to study as a target, and use the information from it to apply to other scenarios. Most people would be familiar with the concept based on medical research: the ‘lab rat’ (or mouse). The common house mouse (Mus musculus) and the brown rat (Rattus norvegicus) are some of the most widely used models for understanding the impact of particular biochemical compounds on physiology and are often used as the testing phase of medical developments before human trials.
A simplified summary of the speciation experiment in Drosophila, starting with a single species and resulting in two reproductively isolated species based on mating and food preference. Source: Ilmari Karonen, adapted from here.
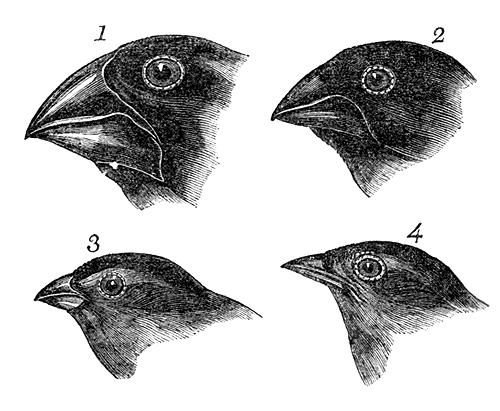
Some of Darwin’s early drawings of the morphological differences in Galapagos finch beaks, which lead to the formulation of the theory of evolution by natural selection.
The sheer diversity of species and form makes African cichlids an ideal model for testing hypotheses and theories about the process of evolution and adaptive radiation. Figure sourced from Brawand et al. (2014) in Nature.
Managing and conserving threatened and endangered species in the wild is a difficult process. There are a large number of possible threats, outcomes, and it’s often not clear which of these (or how many of these) are at play at any one given time. Thankfully, there are also a large number of possible conservation tools that we might be able to use to protect, bolster and restore species at risk.
A diagram of the hierarchy of structure within a species. Remember that ESUs, by definition, should be evolutionary different from one another (i.e. adaptively divergent) whilst MUs are not necessarily divergent to the same degree.
This can lead to a particular paradigm of conservation management: keeping everything separate and pure is ‘best practice’. The logic is that, as these different groups have evolved slightly differently from one another (although there is often a lot of grey area about ‘differently enough’), mixing these groups together is a bad idea. Particularly, this is relevant when we consider translocations (“it’s never acceptable to move an organism from one ESU into another”) and captive breeding programs (“it’s never acceptable to breed two organisms together from different ESUs”). So, why not? Why does it matter if they’re a little different?
Outbreeding depression
Well, the classic reasoning is based on a concept called ‘outbreeding depression’. We’ve mentioned outbreeding depression before, and it is a key concept kept in mind when developing conservation programs. The simplest explanation for outbreeding depression is that evolution, through the strict process of natural selection, has pushed particularly populations to evolve certain genetic variants for a certain selective pressure. These can vary across populations, and it may mean that populations are locally adapted to a specific set of environmental conditions, with the specific set of genetic variants that best allow them to do this.
However, when you mix in the genetic variants that have evolved in a different population, by introducing a foreign individual and allowing them to breed, you essentially ‘tarnish’ the ‘pure’ gene pool of that population with what could be very bad (maladaptive) genes. The hybrid offspring of ‘native’ and this foreign individual will be less adaptive than their ‘pure native’ counterparts, and the overall adaptiveness of the population will decrease as those new variants spread (depending on the number introduced, and how negative those variants are).
An example of how outbreeding depression can affect a species. The original red fish population is not doing well- it is of conservation concern, and has very little genetic diversity (only the blue gene in this example). So, we decide to introduce new genetic diversity by adding in green fish, which have the orange gene. However, the mixture of the two genes and the maladaptive nature of the orange gene actually makes the situation worse, with the offspring showing less fitness than their preceding generations.
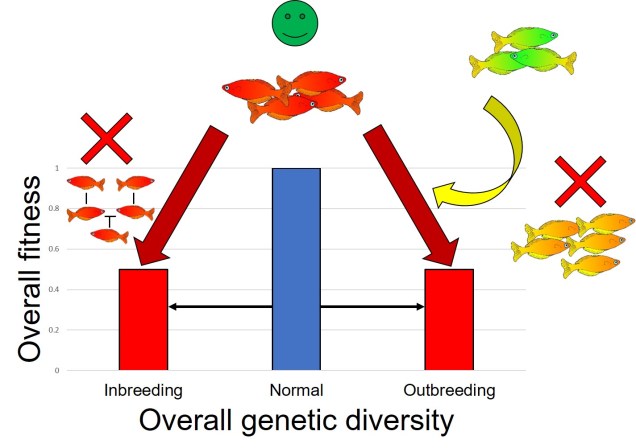
You might be familiar with inbreeding depression, which is based on the loss of genetic diversity from having too similar individuals breeding together to produce very genetically ‘weak’ offspring through inbreeding. Outbreeding depression could be thought of as the opposite extreme; breeding too different individuals introduced too many ‘bad’ alleles into the population, diluting the ‘good’ alleles.
An overly simplistic representation of how inbreeding and outbreeding depression can reduce overall fitness of a species. In inbreeding depression, the lack of genetic diversity due to related individuals breeding with one another makes them at risk of being unable to adapt to new pressures. Contrastingly, adding in new genes from external populations which aren’t fit for the target population can also reduce overall fitness by ‘diluting’ natural, adaptive allele frequencies in the population.
Genetic rescue
It might sound awfully purist to only preserve the local genetic diversity, and to assume that any new variants could be bad and tarnish the gene pool. And, surprisingly enough, this is an area of great debate within conservation genetics.
An example of genetic rescue. This circumstance is identical to the one above, with the key difference being in the fitness of the introduced gene. The orange gene in this example is actually beneficial to the target population: by providing a new, adaptive allele for natural selection to act upon, overall fitness is increased for the red fish population.
The balance
So, what’s the balance between the two? Is introducing new genetic variation a bad idea, and going to lead to outbreeding depression; or a good idea, and lead to genetic rescue? Of course, many of the details surrounding the translocation of new genetic material is important: how different are the populations? How different are the environments (i.e. natural selection) between them? How well will the target population take up new individuals and genes?
Overall, however, the more recent and well-supported conclusion is that fears regarding outbreeding depression are often strongly exaggerated. Bad alleles that have been introduced into a population can be rapidlypurged by natural selection, and the likelihood of a strongly maladaptive allele spreading throughout the population is unlikely. Secondly, given the lack of genetic diversity in the target population, most that need the genetic rescue are so badly maladaptive as it is (due to genetic drift and lack of available adaptive alleles) that introducing new variants is unlikely to make the situation much worse.
An example of how introducing maladaptive alleles might not necessarily lead to decreased fitness. In this example, we again start with our low diversity red fish population, with only one allele (AA). To help boost genetic diversity, we introduce orange fish (with the TT allele) and green fish (with the GG allele) into the population. However, the TT allele is not very adaptive in this new environment, and individuals with the TT gene quickly die out (i.e. be ‘purged’). Individual with the GG gene, however, do well, and continue to integrate into the red population. Over time, these two variants will mix together as the two populations hybridise and overall fitness will increase for the population.
That said, outbreeding depression is not an entirely trivial concept and there are always limitations in genetic rescue procedures. For example, it would be considered a bad idea to mix two different species together and make hybrids, since the difference between two species, compared to two populations, can be a lot stronger and not necessarily a very ‘natural’ process (whereas populations can mix and disjoin relatively regularly).
The reality of conservation management
Conservation science is, at its core, a crisis discipline. It exists solely as an emergency response to the rapid extinction of species and loss of biodiversity across the globe. The time spent trying to evaluate the risk of outbreeding depression – instead of immediately developing genetic rescue programs – can cause species to tick over to the afterlife before we get a clear answer. Although careful consideration and analysis is a requirement of any good conservation program, preventing action due to almost paranoid fear is not a luxury endangered species can afford.
The classic way for new genetic variants to appear is often thought of as mutation: changes in a single base in the DNA are caused by various external processes such as chemical, physical or environmental influences (such as the sci-fi classics like UV rays or toxic chemicals). Although these forms of mutations happen very rarely and certainly don’t have the same effects comic books would leave you to believe, new mutations can occur relatively rapidly depending on the characteristics of the species. However, the most common way for new mutations to occur is actually part of the DNA replication process: copying DNA is not always perfect and even though the relevant proteins essentially run a spellcheck, sometimes the copy is not 100% perfect and new mutations occur.
An example of how adaptation can occur from a new mutation. In this example, we have one gene (TTXTT), with initial only one allele (variant), TTATT. In the second generation (row), a mutation occurs in one individual which creates a new, second allele: TTGTT. This allele is favoured over the TTATT allele, and in the next generation it’s frequency increases as the alternative allele frequency decreases (the pattern is shown in the frequency values on the right side).
Alternatively, genetic variation might already be present within a species or population. This is more likely if population sizes are large and populations are well connected and interbreeding. We refer to this diverse initial gene pool as ‘standing genetic variation’: that is, the amount of genetic variation within the population or species before the selective pressure requiring adaptation. Standing genetic variation can be thought of as the ‘diversity of choices’ for natural selection to act upon: the variants are readily available, and if a good choice exists it will be favoured by natural selection and become more widespread within the population or species (i.e. evolve).
A slightly more complex example of how adaptation can occur from standing variation, this time with two different genes. One codes for fur colour, with two different alleles: GCATA codes for orange fur, and GCGTA codes for grey fur. The other gene codes for ear tufts, with TTCCT coding for tufts and TCCCT coding for no tufts. Natural selection favours both orange fur and tufted ears, and cats with these traits reproduce more frequently than those without (see graph below). These cats probably look familiar.The frequency of all four alleles (i.e. either allele for both genes) over the generations in the above figure. Clearly, we can see how adaptation rapidly favours orange fur and tufted ears over grey fur and non-tufted ears with the shifts in frequencies over the different alleles.
We’ve discussed standing genetic variation before on The G-CAT, but often in a different light (and phrasing). For example, when we’ve talked about founder effect: that is, when a population is formed from only a few different individuals which causes it to be very genetically depauperate. In populations under strong founder effect, there is very little standing genetic variation for natural selection to act upon. This has long been an enigma for many pest species: how have they managed to proliferate so widely when they often originate from so few individuals and lack genetic diversity?
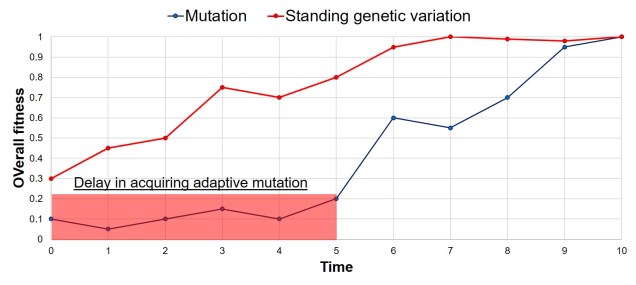
A rough example of the speed of adaptation depending on how the adaptive allele originated: whether it was already present (in the form of standing variation), or whether it was created by a new mutation. As one would expect, there is a significant lag delay in adaptation in the mutation scenario, based on the time it takes for said adaptive mutation to be created through relatively random processes. Thus, a positively selected allele from standing variation can allow a species to adapt much faster than waiting for a positive mutation to occur.
An extreme example of alternate splicing of one gene. We start with a single gene, composed of 5 (A–E) main gene elements (exons). Different environmental pressures (like fire risk, flooding, cold weather or predators, for example) cause the organism to use different combinations of these exons to make different proteins (right side; A–D). Actual alternate splicing is not usually this straight-forward (one gene doesn’t conveniently split into four forms depending on the threat), but the process is generally the same.