Unravelling the evolutionary history of organisms – one of the main goals of phylogenetic research – remains a challenging prospect due to a number of theoretical and analytical aspects. Particularly, trying to reconstruct evolutionary patterns based on current genetic data (the most common way phylogenetic trees are estimated) is prone to the erroneous influence of some secondary factors. One of these is referred to as ‘incomplete lineage sorting’, which can have a major effect on how phylogenetic relationships are estimated and the statistical confidence we may have around these patterns. Today, we’re going to take a look at incomplete lineage sorting (shortened to ILS for brevity herein) using a game-based analogy – a Pachinko machine. Or, if you’d rather, the same general analogy also works for those creepy clown carnival games, but I prefer the less frightening alternative.
It’s been a few minutes (okay, several weeks) since the last post here on The G-CAT. Naturally, over that time I’ve spent holidays with both my own family and my partner’s family. Hopefully, you’ve enjoyed your own Christmas/New Year/Other-Non-Denominational-Celebrations break (and for us Aussies, that you’ve managed to avoid much of the devastation of the recent bushfire epidemic).
Because of this period of time (and a few other more pressing deadlines I had for the start of the new year), I haven’t prepared a new post in some time. However, I’d like to take this time to address how the nature of this blog might change of the next year or so (and into the future).
A new schedule
For those of you who keep more up-to-date with my academic progress, you’ll be aware that this year is the final year of my PhD. As it stands, I’m due to submit my thesis in August of this year (which feels much, much sooner than it really is). Similarly, for anyone who has ever interacted with a PhD student in the final year of their studies, you’ll also be aware that this can be a time of high stress, stacking deadlines and the overall impending doom of D-Day (thesis submission).
In light of all of this, I have decided to move away from the more predictable fortnightly post routine in favour of a more organic timetable. This will likely mean a fairly significant reduction in the frequency of blog posts, whereby I will post as a topic comes into my mind or when it appears relevant to other parts of my studies (e.g. in reading for writing manuscripts, etc.). This decision has also been playing on my mind for some time to also balance the quality of the posts I write: in some circumstances, I feel like the consistent deadline of once per fortnight causes some posts to suffer a little as I rush to produce something at least in the vicinity of every second Wednesday.
The future of The G-CAT
It is not my intention to completely abandon this project: The G-CAT is something that I have invested a fair time and inspiration into and provides a solid avenue for science communication. As always, my inbox (on whichever platform you choose) is wide open for suggestions on topics of discussion. I’m looking forward to a more organic schedule that will allow me to properly explore and expand on the topics of interest whilst maintaining a healthy balance of PhD progression and down-time.
One very effective summary statistic that we might choose to use is the site-frequency spectrum (aka the allele frequency spectrum). Not to be confused with other measures of allele frequency which we’ve discussed before (like Fst), the site-frequency spectrum (abbreviated to SFS) is essentially a histogram of how frequent certain alleles are within our dataset. To do this, the SFS classifies each allele into a certain category based on how common it is, tallying up the number of alleles that occur at that frequency. The total number of categories would be the maximum number of possible alleles: for organisms with two copies of every chromosome (‘diploids’, including humans), this means that there are double the number of samples included. For example, a dataset comprised of genomic sequence for 5 people would have 10 different frequency bins.
An example of the 1DSFS for a single population, taken from a real dataset from my PhD. Left: the full site-frequency spectrum, counting how many alleles (y-axis) occur a certain number of times (categories of the x-axis) within the population. In this example, as in most species, the vast majority of our DNA sequence is non-variable (frequency = 0). Given the huge disparity in number of non-variable sites, we often select on the variable ones (and even then, often discard the 1 category to remove potential sequencing errors) and get a graph more like the right. Right: the ‘realistic’ 1DSFS for the population, showing a general exponential decline (the blue trendline) for the more frequent classes. This is pretty standard for an SFS. ‘Singleton’ and ‘doubleton’ are alternative names for ‘alleles which occur once’ and ‘alleles which occur twice’ in an SFS.
Expanding the SFS to multiple populations
Further to this, we can expand the site-frequency spectrum to compare across populations. Instead of having a simple 1-dimensional frequency distribution, for a pair of populations we can have a grid. This grid specifies how often a particular allele occurs at a certain frequency in Population A and at a different frequency in Population B. This can also be visualised quite easily, albeit as a heatmap instead. We refer to this as the 2-dimensional SFS (2DSFS).
An example of a 2DSFS, also taken from my PhD research. In this example, we are comparing Population A, containing 5 individuals (as diploid, 2 x 5 = max. of 10 occurrences of an allele) with Population B, containing 4 individuals. Each row denotes the frequency at which a certain allele occurs in Population B whilst the columns indicate the frequency a certain allele occurs in Population A. Each cell therefore indicates the number of alleles that occur at the exact frequency of the corresponding row and column. For example, the first cell (highlighted in green) indicates the number of alleles which are not found in either Population A or Population B (this dataset is a subsample from a larger one). The yellow cell indicates the number of alleles which occur 4 times in Population B and also 4 times in Population A. This could mean that in one of those Populations 4 individuals have one copy of that allele each, or two individuals have two copies of that allele, or that one has two copies and two have one copy. The exact composition of how the alleles are spread across samples within each population doesn’t matter to the overall SFS.
The same concept can be expanded to even more populations, although this gets harder to represent visually. Essentially, we end up with a set of different matrices which describe the frequency of certain alleles across all of our populations, merging them together into the joint SFS. For example, a joint SFS of 4 populations would consist of 6 (4 x 4 total comparisons – 4 self-comparisons, then halved to remove duplicate comparisons) 2D SFSs all combined together. To make sense of this, check out the diagrammatic tables below.
A summary of the different combinations of 2DSFSs that make up a joint SFS matrix. In this example we have 4 different populations (as described in the above text). Red cells denote comparisons between a population and itself – which is effectively redundant. Green cells contain the actual 2D comparisons that would be used to build the joint SFS: the blue cells show the same comparisons but in mirrored order, and are thus redundant as well.
Expanding the above jSFS matrix to the actual data, this matrix demonstrates how the matrix is actually a collection of multiple 2DSFSs. In this matrix, one particular cell demonstrates the number of alleles which occur at frequency xin one population and frequency yin another. For example, if we took the cell in the third row from the top and the fourth column from the left, we would be looking at the number of alleles which occur twice in Population B and three times in Population A. The colour of this cell is moreorless orange, indicating that ~50 alleles occur at this combination of frequencies. As you may notice, many population pairs show similar patterns, except for the Population C vs Population D comparison.
The different forms of the SFS
Which alleles we choose to use within our SFS is particularly important. If we don’t have a lot of information about the genomics or evolutionary history of our study species, we might choose to use the minor allele frequency (MAF). Given that SNPs tend to be biallelic, for any given locus we could have Allele A or Allele B. The MAF chooses the least frequent of these two within the dataset and uses that in the summary SFS: since the other allele’s frequency would just be 2N – the frequency of the other allele, it’s not included in the summary. An SFS made of the MAF is also referred to as the folded SFS.
Alternatively, if we know some things about the genetic history of our study species, we might be able to divide Allele A and Allele B into derived or ancestral alleles. Since SNPs often occur as mutations at a single site in the DNA, one allele at the given site is the new mutation (the derived allele) whilst the other is the ‘original’ (the ancestral allele). Typically, we would use the derived allele frequency to construct the SFS, since under coalescent theory we’re trying to simulate that mutation event. An SFS made of the derived alleles only is also referred to as the unfolded SFS.
Applications of the SFS
How can we use the SFS? Well, it can moreorless be used as a summary of genetic variation for many types of coalescent-based analyses. This means we can make inferences of demographic history (see here for more detailed explanation of that) without simulating large and complex genetic sequences and instead use the SFS. Comparing our observed SFS to a simulated scenario of a bottleneck and comparing the expected SFS allows us to estimate the likelihood of that scenario.
A representative example of how a bottleneck causes a shift in the SFS, based on a figure from a previous post on the coalescent. Centre: the diagram of alleles through time, with rarer variants (yellow and navy) being lost during the bottleneck but more common variants surviving (red). Left: this trend is reflected in the coalescent trees for these alleles, with red crosses indicating the complete loss of that allele. Right: the SFS from before (in red) and after (in blue) the bottleneck event for the alleles depicted. Before the bottleneck, variants are spread in the usual exponential shape: afterwards, however, a disproportionate loss of the rarer variants causes the distribution to flatten. Typically, the SFS would be built from more alleles than shown here, and extend much further.
A similar diagram as above, but this time with an expansion event rather than a bottleneck. The expansion of the population, and subsequent increase in Ne, facilitates the mutation of new alleles from genetic drift (or reduced loss of alleles from drift), causing more new (and thus rare) alleles to appear. This is shown by both the coalescent tree (left) and a shift in the SFS (right).
The SFS can even be used to detect alleles under natural selection. For strongly selected parts of the genome, alleles should occur at either high (if positively selected) or low (if negatively selected) frequency, with a deficit of more intermediate frequencies.
Adding to the analytical toolbox
The SFS is just one of many tools we can use to investigate the demographic history of populations and species. Using a combination of genomic technologies, coalescent theory and more robust analytical methods, the SFS appears to be poised to tackle more nuanced and complex questions of the evolutionary history of life on Earth.
Meaning: Octorokus from [octorok] in Hylian; infletus from [inflate] in Latin.
Translation: inflating octorok; all varieties use an inflatable air sac derived from the swim bladder to float and scan the horizon.
Varieties
Octorokus infletus hydros [aquatic morphotype]
Octorokus infletus petram [mountain morphotype]
Octorokus infletus silva [forest morphotype]
Octorokus infletus arctus [snow morphotype]
Octorokus infletus imitor [deceptive morphotype]
The various morphotypes of inflating octoroks. A: The water octorok, considered the morphotype closest to the ancestral physiology of the species. B: The forest octorok, with grass camouflage. C: The deceptive octorok, which has replaced its tufted vegetation with a glittering chest as bait. D: The mountainous octorok, with rock camouflage. E: The snow octorok, with tundra grass camouflage.
Common name
Variable octorok
Taxonomic status
Kingdom Animalia; Phylum Mollusca; Class Cephalapoda; Order Octopoda; Family Octopididae; GenusOctorokus; Speciesinfletus
Conservation status
Least Concern
Distribution
The species is found throughout all major habitat regions of Hyrule, with localised morphotypes found within specific habitats. The only major region where the variable octorok is not found is within the Gerudo Desert, suggesting some remnant dependency of standing water.
The region of Hyrule, with the distribution of octoroks in blue. The only major region where they are not found is the Gerudo Desert in the bottom left.
Habitat
Habitat choice depends on the physiology of the morphotype; so long as the environment allows the octorok to blend in, it is highly likely there are many around (i.e. unseen).
Behaviour and ecology
The variable octorok is arguably one of the most diverse species within modern Hyrule, exhibiting a large number of different morphotypic forms and occurring in almost all major habitat zones. Historical data suggests that the water octorok (Octorokus infletus hydros) is the most ancestral morphotype, with ancient literature frequently referring to them as sea-bearing or river-traversing organisms. Estimates from the literature suggests that their adaptation to land-based living is a recent evolutionary step which facilitated rapid morphological radiation of the lineage.
Several physiological characteristics unite the variable morphological forms of the octorok into a single identifiable species. Other than the typical body structure of an octopod (eight legs, largely soft body with an elongated mantle region), the primary diagnostic trait of the octorok is the presence of a large ‘balloon’ with the top of the mantle. This appears to be derived from the swim bladder of the ancestral octorok, which has shifted to the cranial region. The octorok can inflate this balloon using air pumped through the gills, filling it and lifting the octorok into the air. All morphotypes use this to scan the surrounding region to identify prey items, including attacking people if aggravated.
A water morphotype octorok with balloon inflated.
Diets of the octorok vary depending on the morphotype and based on the ecological habitat; adaptations to different ecological niches is facilitated by a diverse and generalist diet.
Demography
Although limited information is available on the amount of gene flow and population connectivity between different morphotypes, by sheer numbers alone it would appear the variable octorok is highly abundant. Some records of interactions between morphotypes (such as at the water’s edge within forested areas) implies that the different types are not reproductively isolated and can form hybrids: how this impacts resultant hybrid morphotypes and development is unknown. However, given the propensity of morphotypes to be largely limited to their adaptive habitats, it would seem reasonable to assume that some level of population structure is present across types.
Adaptive traits
The variable octorok appears remarkably diverse in physiology, although the recent nature of their divergence and the observed interactions between morphological types suggests that they are not reproductively isolated. Whether these are the result of phenotypic plasticity, and environmental pressures are responsible for associated physiological changes to different environments, or genetically coded at early stages of development is unknown due to the cryptic nature of octorok spawning.
All octoroks employ strong behavioural and physiological traits for camouflage and ambush predation. Vegetation is usually placed on the top of the cranium of all morphotypes, with the exact species of plant used dependent on the environment (e.g. forest morphotypes will use grasses or ferns, whilst mountain morphotypes will use rocky boulders). The octorok will then dig beneath the surface until just the vegetation is showing, effectively blending in with the environment and only occasionally choosing to surface by using the balloon. Whether this behaviour is passed down genetically or taught from parents is unclear.
Management actions
Few management actions are recommended for this highly abundant species. However, further research is needed to better understand the highly variable nature and the process of evolution underpinning their diverse morphology. Whether morphotypes are genetically hardwired by inheritance of determinant genes, or whether alterations in gene expression caused by the environmental context of octoroks (i.e. phenotypic plasticity) provides an intriguing avenue of insight into the evolution of Hylian fauna.
Nevertheless, the transition from the marine environment onto the terrestrial landscape appears to be a significant stepping stone in the radiation of morphological structures within the species. How this has been facilitated by the genetic architecture of the octorok is a mystery.
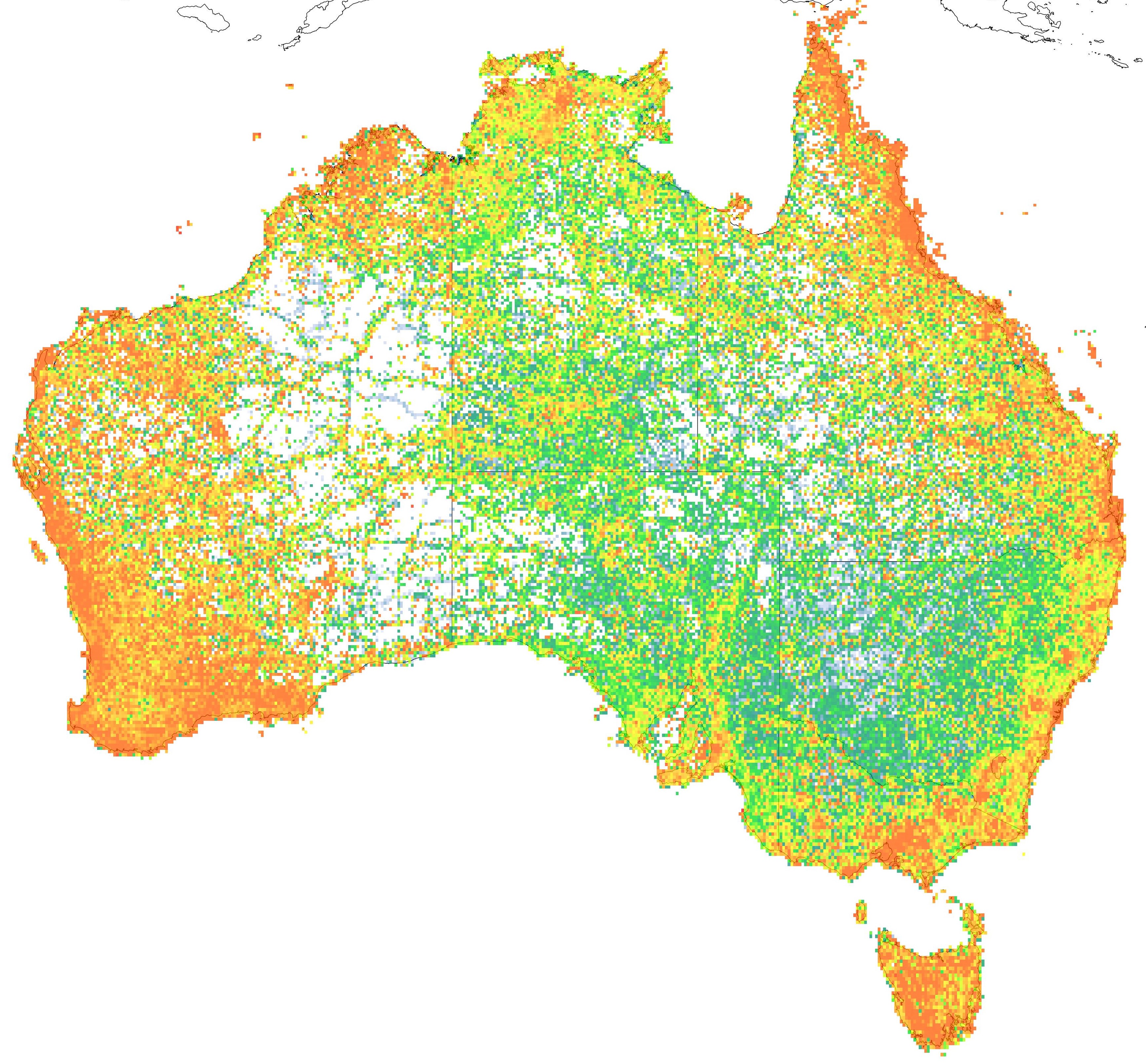
A map representing the relative proportion of endemic species in Australia, generated through the Atlas of Living Australia. The colours range from no (white; 0% endemics) or little (blue) to high levels of endemism (red; 100% of species are endemic). As you can see, some biogeographic hotspots are clearly indicated (southwest WA, the east coast, the Kimberley ranges).
But the real question is: why are there so many endemics in Australia? What is so special about our country that lends to our unique flora and fauna? Although we naturally associate tropical regions with lush, vibrant and diverse life, most of Australia is complete desert. That said, most of our species are concentrated in the tropical regions of the country, particularly in the upper east coast and far north (the ‘Top End’).
There are a number of different factors which contribute to the high species diversity of Australia. Most notably is how isolated we are as a continent: Australia has been separated from most of the rest of the world for millions of years. In this time, the climate has varied dramatically as the island shifted northward, creating a variety of changing environments and unique ecological niches for species to specialise into. We refer to these species groups as ‘Gondwana relicts’, since their last ancestor with the rest of the world would have been distributed across the supercontinent Gondwana over 100 million years ago. These include marsupials, many birds groups (including ratites and megapodes), many fish groups and a plethora of others. A Gondwanan origin explains why they are only found within Australia, southern Africa and South America (the closest landmass that was also historically connected to Gondwana).
Early arrivals and naturalisation to the Australian ecosystem
Eventually, this connection also brought with them one of our most iconic species; the dingo. Estimates of their arrival dates the migration at around 6 thousand years ago. As Australia’s only ‘native’ dog, there has been much debate about its status as an Australian icon. To call the dingo ‘native’ implies it’s always been there: but 6 thousand years is more than enough time to become ingrained within the ecosystem in a stable fashion. So, to balance the debate (and prevent the dingo from being labelled as an ‘invasive pest’ unfairly), we often refer to them as ‘naturalised’. This term helps us to disentangle modern-day pests, many of which our immensely destructive to the natural environment, from other species that have naturally migrated and integrated many years ago.
Although it may not be a “true native”, the dingo will forever be a badge of our native species pride.
Invaders of the Australian continent
Of course, we can never ignore the direct impacts of humans on the ecosystem. Particularly with European settlement, another plethora of animals were introduced for the first time into Australia; these were predominantly livestock animals or hunting-related species (both as predators and prey). This includes the cane toad, widely regarded as one of the biggest errors in pest control on the planet.
When European settlers in the 1930s attempted to grow sugar cane in the far eastern part of the country, they found their crops decimated by a local beetle. In an effort to eradicate them, they brought over a species of cane toad, with the idea that they would control the beetle population and all would be well. Only, cane toads are particularly lazy and instead of targeting the cane beetles, they just thrived on all the other native invertebrates around. They’re also very resilient and adaptable (and highly toxic), so their numbers exploded and they’ve since spread across a large swathe of the country. Their toxic skin makes them fatal food objects for many native predators and they strongly compete against other similar native animals (such as our own amphibians). The cane toad introduction of 1935 is the poster child of how bad failed pest control can be.
This guy here, he’s a bastard. Spotted in my parent’s backyard in Ipswich, QLD. Source: me, with spite.
But is native always better?
History tells a very stark tale about the poor native animals and the ravenous, rampaging pest species. Because of this, it is a widely adopted philosophical viewpoint that ‘native is always best’. And while I don’t disagree with the sentiment (of course we need to preserve our native wildlife, and not the massively overabundant pests), there are rare examples where nature is a little more complicated. In Australia, this is exemplified in the noisy miner.
The noisy miner is a small bird which, much like its name implies, is incredibly noisy and aggressive. It’s highly abundant, found predominantly throughout urban and suburban areas, and seems to dominate the habitat. It does this by bullying out other bird species from nesting grounds, creating a monopoly on the resource to the exclusion of many other species (even larger ones such as crows and magpies). Despite being native, it seems to have thrived on human alteration of the landscape and is a serious threat to the survival and longevity of many other species. If we thought of it solely under the ‘nature is best’ paradigm, we would dismiss the noisy miner as ‘doing what it should be.’ The truth is really more of a philosophical debate: is it natural to let the noisy miner outcompete many other natives, possibly resulting in their extinction? Or is it only because of human interference (and thus is our responsibility to fix) that the noisy miner is doing so well in the first place? It’s not a simple question to answer, although the latter seems to be incredibly important.
An example of the aggressive behaviour of the noisy miner (top), swooping down on a pied currawong (bottom). Despite the size differences, noisy miners will frequently attempt to harass and scare off other larger birds. Image source: Bird Ecology Study Group website.
The amazing biodiversity of Australia is a badge of honour we should wear with patriotic pride. Conservation efforts of our endemic fauna are severely limited by a lack of funding and resources, and despite a general acceptance of the importance of diverse ecosystems we remain relatively ineffective at preserving it. Understanding and connecting with our native wildlife, whilst finding methods to control invasive species, is key to conserving our wonderful ecosystems.