The environmental context of adaptation
We’ve talked many times before about how species evolve in response to some kind of environmental pressure, which favours (or disfavours) certain traits within that species. Over time, this drives changes in the frequencies of species traits and alters the overall average phenotype of that species (sometimes slowly, sometimes rapidly).
While we usually talk about the environment in terms of abiotic conditions such as temperature or climate, biotic factors are equally important: that is, the parts of the environment which are themselves also alive. Because of this, changes in one species can have profound repercussions on other species linked within the ecosystem. Thus, the evolution of one species is intrinsically linked to the evolution of other relevant species within the ecosystem: often, these connected evolutionary pathways battle with one another as each one changes. Let’s take a look at a few different examples of how evolution of one species may impact the evolution of another.
Predator-prey coevolution
One of the most obvious ways the evolution of two different species can interact is in predator and prey relationships. Naturally, prey species evolve to be able to defend themselves from predators in various ways, such as crypsis (e.g. camouflage), toxicity or behavioural changes (such as nocturnalism or group herding). Contrastingly, predators will evolve new and improved methods for detecting and hunting prey, such as enhanced senses, venom and stealth (through soft-padded feet, for example).
There are millions of possible examples of predator-prey coevolution that could be used as examples here, based on the continual drive for one species to get the upper hand over the other. But one that comes to mind is of a creature that I learnt about while on holiday in Scandinavia: the pine marten, and how it affects squirrels.

The pine marten is a species in the mustelid family, along with otters, weasels, stoats, and wolverines. Like many mustelids, they are carnivorous mammals which feed on a variety of different prey items like rodents, small birds and insects. One of the more abundant species that they prey upon are squirrels: both red squirrels and grey squirrels are potential food for the cute yet savage pine marten.
However, within the distribution of pine martens (across much of Europe), red squirrels are the native species and grey squirrels are invasive, originating from North America. Because of the long-lasting relationship between red squirrels and pine martens, they’ve co-evolved: most notably, by red squirrels changing to a mostly arboreal lifestyle and avoiding the ground as much as possible. Grey squirrels, however, have not had the evolutionary history to learn this lesson and are easy food for a smart pine marten. Thus, in regions where pine martens have been conserved or reintroduced, they are actively controlling the invasive grey squirrel population, which in turn boosts the native red squirrel population by reduction of competition. The coevolutionary link between red squirrels and pine martens is critical for combating the invasive species.


Host-parasite coevolution
In a similar vein to predator and prey coevolution, pathogenic species and their unfortunate hosts also undergo a sort of ‘arms race’. Parasites must keep evolving new ways to infect and transmit to hosts as the hosts evolve new methods of resisting and avoiding the infecting species. This spiralling battle of evolutionary forces is dubbed as the ‘Red Queen hypothesis’, formulated in 1973 by Leigh Van Valen and used to describe many other forms of coevolution. The name comes from Lewis Carroll’s Through the Looking Glass, and one quote in particular:
‘Now, here, you see, it takes all the running you can do, to keep in the same place’.
The quote references how species must continually adapt and respond to the evolution of other species just keep existing and prevent extinction. Species that remain static and stop evolving will inevitably go extinct as the world around them changes.
Mimicry
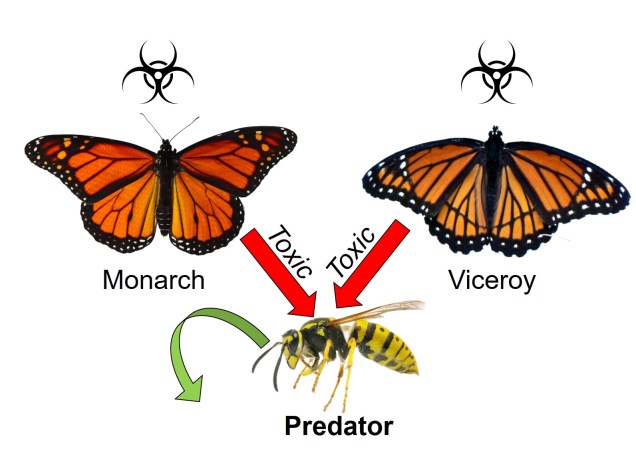
Plenty of other strange and unique mechanisms of coevolution exist within nature. One of them is mimicry, the process by which one species attempts to look like another to protect itself. The most iconic group known for this is butterflies: many species, although they may be evolutionarily very different, share similar colouration patterns and body shapes as mimics. Depending on the nature of the copy, mimicry can be classified into two broad categories. In either case, the initial ‘reference’ species is toxic or unpalatable to predators and uses a type of colour signal to communicate this: think of the bright yellow colours of bees and wasps or the red of ladybirds. Where the two categories change is in the nature of the ‘mimic’ species.
Müllerian mimicry
If the mimic is also toxic or unpalatable, we call this Müllerian mimicry (after Johann Friedrich Theodor Müller). By sharing the same colouration patterns and both being toxic, the two mimicking species boost the potential for the signal to be learnt by predators. If a predator eats either species, it will associate that colour pattern with toxicity and neither species are as likely to be preyed upon in the future. In this sense, it is a cooperative coevolutionary relationship between the two physically similar species.
Batesian mimicry
In contrast, the mimic might not actually be toxic or unpalatable, and simply copying a toxic species. This is referred to as Batesian mimicry (after Henry Walter Bates), and involves a mimic species relying on the association of colour and toxicity to have been learnt by predators through the ‘reference’ species. Although the mimic is not toxic, it is essentially piggy-backing on the hard evolutionary work that has already been done by the actually toxic species. In this case, the coevolutionary relationship is more parasitic as the mimic benefits from the ‘reference’ but the favour is not returned.

Coevolution of species and the importance of species interactions
There are countless of other species interactions which could drive coevolutionary relationships in nature. These can include various forms of symbiosis, or the response of different species to ecosystem engineers: that is, species that can change and shape the environment around them (such as corals in reef systems). Understanding how a species evolves within its environment thus needs to consider how many other local species are also evolving and responding in their own ways.