Adaptation and natural selection
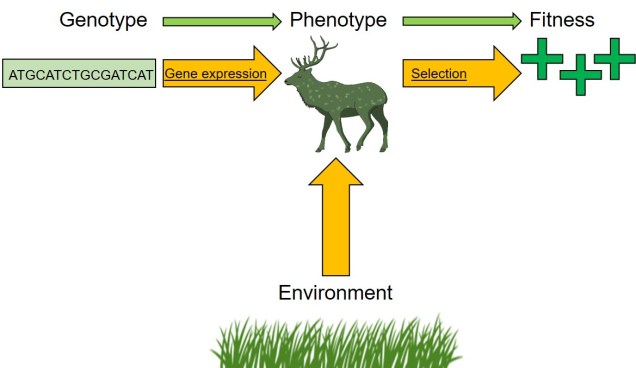
Adaptation via natural selection is one of the most fundamental components of understanding evolution. It describes how species can continually evolve new, innovative traits and produce the wondrous diversity of the natural world. This process is inevitably underpinned by particular heritable traits often linked to particular genetic variants (alleles). Remember that the underlying genetic trait (the allele) is referred to as the genotype; the physical outcomes of that allele (i.e. how it changes the physiological, behaviour or ecology of the organism) is the phenotype; and the scale of the benefit of possessing that trait is referred to as its fitness. Under the normal process of natural selection, phenotypes which increase fitness are selected for, which results in a shift in genotypes underpinning it.
Maladaptation
Of course, however, where there’s good there’s also bad, and not all traits are favoured by natural selection. Under the same process that can cause alleles which relate to adaptive traits to increase in frequency, the inverse may happen. That is, alleles (particularly new mutations) which relate to negative traits – such as those that disrupt the structure of a particularly important proteins – may be selected against by natural selection if they lead to reduced fitness. The balance of ‘adaptiveness’, and whether a trait is selected for or against (and how strong that selection is) is directly related to the overall fitness value of that trait.
As with many things in biology (see also: species concepts), there are a plethora of ways to describe ‘adaptation’ based on the way it is measured and whether you are referring to a noun (i.e. the trait is an adaptation) or the process (i.e. the trait evolves by adaptation). Just to be confusing.
The flipside of that means that the ways to describe the inverse process – commonly referred to as maladaptation – are similarly diverse. These can include the mere absence of a trait known to be adaptive; or the process itself driving the declination of fitness; or the general state of a population having low fitness. Regardless, each of these terms ultimately lead to a relatively singular concept – the idea that some traits are unfavoured by natural selection and should (evolution willing) be purged from the pool of traits.

How does maladaptation occur?
Much like adaptation, there are a number of different processes which may drive the formation or accumulation of maladaptive traits within a population or species. Generally, any process that typically hinders or limits the capacity for adaptive evolutionary processes, such as population fragmentation, inbreeding depression or even artificial selection, may drive maladaptation. These might prevent the accumulation or maintenance of adaptive genetic diversity, which when removed from the gene pool confers maladaptive traits through the loss of adaptive potential (definition 1).
For example, significant inbreeding depression can result in increased genetic load through the inability of populations to ‘purge’ maladaptive alleles from the gene pool (typically through recombination across different loci). Another way this can lead to maladaptation (definition 3) is when the lack of adaptive alternatives results in the ‘most adaptive’ phenotype being selected over others in a “best of the worst”-type scenario. This where the distinction between relative fitness (i.e. the fitness of the allele relative to other available alleles) and absolute fitness (i.e. the total fitness benefit that allele confers to the population) is important. In dire situations, even the most relatively fit trait might still have low absolute fitness (if we could compare to a non-impacted population as a reference).
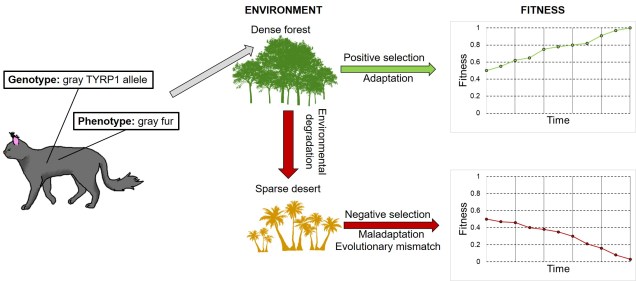
Evolutionary mismatch
Of course, there are a number of other ways maladaptation can occur. One of these is dubbed ‘evolutionary mismatch’, and is the result of how a given trait and the environment interact to create fitness. Remember that the adaptiveness of a trait is a function of both the trait itself and the environment it occurs in. Evolutionary mismatch can happen when a particular trait that was previously adaptive under a certain selective environment is placed into a new environment, under which that trait is no longer adaptive (or has an associated fitness penalty, even). For example, consider how adaptive a given fur coat colour might be in different environments for a predator (see below).
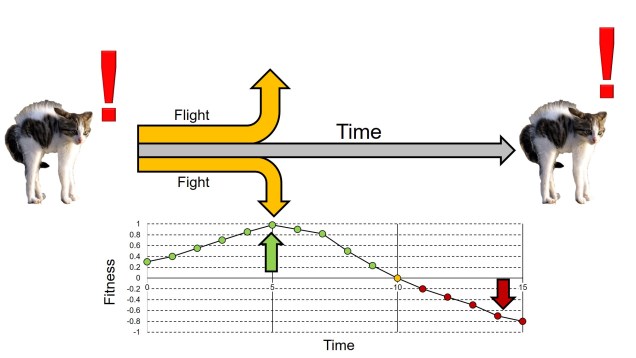
Adaptive trade-offs
Given the complex nature of phenotypes, other maladaptations might result from trade-offs in short-term benefits and long-term costs (or vice versa). A classic example of this is the ‘fight-or-flight’ response strategy, whereby an individual has an immediate physiological response (predominantly hormonal) to a given stressor which instinctively prepares them to engage or flee from the threat. In the short-term, this response is highly adaptive, as the ability to rapidly respond to a potentially life-or-death scenario can, understandably, be life-saving. As anyone dealing with anxiety can tell you, however, continual exposure to this response (particularly when the ‘perceived’ threat is more substantial than the real threat) can lead to significant fitness costs stemming from high blood pressure, expenditure of nervous energy and impaired immune response.
Pleiotropy
One other aspect we’ve touched on before is the notion of pleiotropy: that is, that a single gene may be linked to a number of different phenotypes based on its function. For seemingly complex organisms, the linkage of these different traits – each of which may be conferring different levels of fitness – represents a type of adaptive trade-off as well. ‘Maladaptive’ traits might be maintained if they are linked to a series of other adaptive traits which, in sum, lead to higher fitness.

What are the consequences of maladaptation?
As one might expect, there are a number of consequences of maladaptation. Most directly, this relates to a direct loss of fitness in the population or species, mostly commonly through reduced survivability of individuals or reproductive success. As a flow-on effect, these factors can lead to reduced genetic diversity in future generations, and by proxy the adaptive potential of the population. Without alternative genetic variation to provide adaptive benefits, large-scale and long-lasting maladaptation may initiate (or contribute to) extinction vortex dynamics, providing a scenario of high extinction risk that is difficult to escape by natural means.
Naturally, this is a scenario of particular concern for species already threatened by other means. The rapidly shifting environment, both in terms of the global contemporary climate but also other human-induced habitat modifications, may shift the ‘adaptive goalposts’ for a swathe of species, pushing their phenotypes away from adaptive peaks (and potentially others in the opposite direction). How species respond to the Anthropocene is critical to determine the future of global biodiversity, and considering maladaptation is a key component for understanding how best to conserve our biota.

2 thoughts on “The Bad and the Ugly of evolution: an introduction to maladaptation”